1. Introduction: Development is How You Became You
Many of the processes we’ve been discussing in these tutorials about eukaryotic gene regulation play a key role during embryonic development. During this early phase in life, the body changes in size and form from a fertilized egg into a complex multicellular organism. As you’ll see in the cards below, the biological story of how you became you is quite astonishing.
[qwiz summary=”false” qrecord_id=”sciencemusicvideosMeister1961-The Story of You (Development)(2.0)”]
[h]The story of you
[i]Biohaiku
Everyone’s story:
From a single zygote to
Vast complexity
[q]Your existence, as a distinct genetic being, began when your father’s sperm fertilized your mother’s egg.
[q] Fertilization converted you into a diploid zygote. It also activated the formerly metabolically dormant egg…an egg that had been stored in your mother’s ovaries since she was an early embryo.
[q]Following fertilization you underwent a period of rapid cell division called cleavage. Note that the images below show this process occurring in a sea urchin, an organism often used in studies of development.
As your cells divided, they also migrated, positioning themselves in such a way as to create a fluid-filled ball of cells (3 and 4).
[q] After a few weeks of continued development involving processes such as cell division, programmed cell death, and gene activation leading to cellular differentiation, you looked something like this:
[q] A few decades later, you might find yourself looking like this!
[/qwiz]
We cover the topic of development in detail in our college curriculum. As an AP Biology student, here’s what you need to know.
2. (Almost) All Cells in an Organism are Genomically Equivalent
We’ve mentioned this key principle before, but it bears repeating. In any multicellular organism, with just a few exceptions, all the cells have the same DNA. In other words, the cells in your body are genomically equivalent. The cells that make up the lens of your eye have the same DNA as the cells that produce your toenails. The differences between cells are related to differences in gene expression, not to differences in the cells’ genetic material.
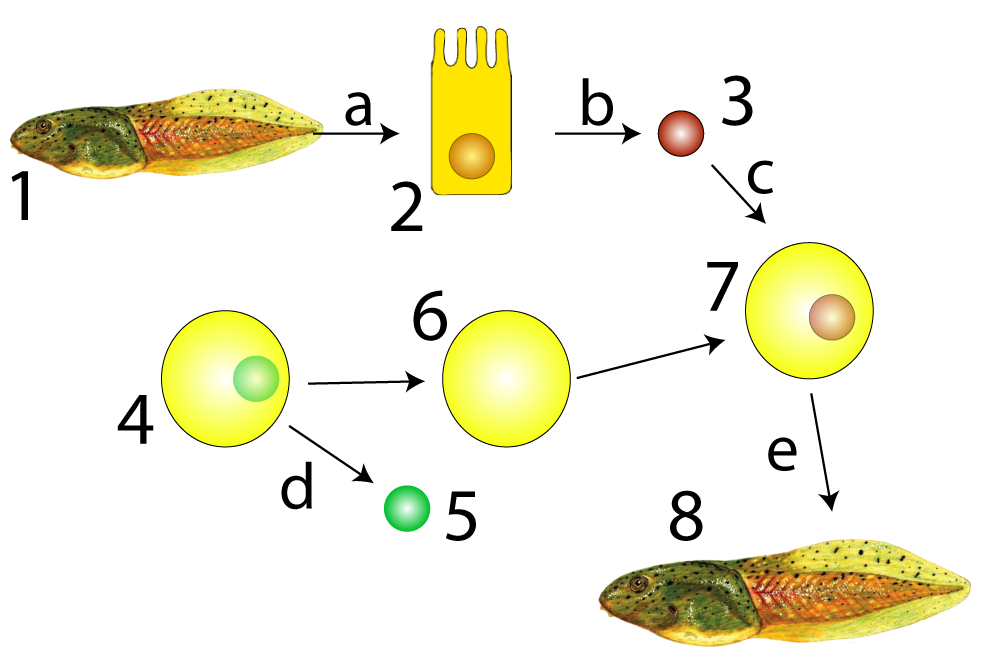
Biologists’ ability to clone animals through somatic cell nuclear transfer is proof of this principle. Cloning is a kind of asexual reproduction in which a somatic cell provides the genetic material from which an entire new organism is formed. A clone is an exact genetic copy of the organism from which it was cloned. You can think of a clone as a younger identical twin with the same exact genes.
Sir John Gurdon, the recipient of the 2012 Nobel Prize for Physiology or Medicine, carried out the experiment shown at left in the late 1950s. He extracted an intestinal cell (2) from a tadpole (1) and then extracted the nucleus from that intestinal cell (3). He separately took a frog egg cell (4) and removed its nucleus (5) creating an enucleated egg (6). Enucleated means that this egg cell had had its nucleus (and therefore its genetic material) removed. Gurdon then placed the intestinal cell’s nucleus into the enucleated frog egg cell (shown at 7). This egg cell was able to develop into a new tadpole: a genetic clone of tadpole number 1.
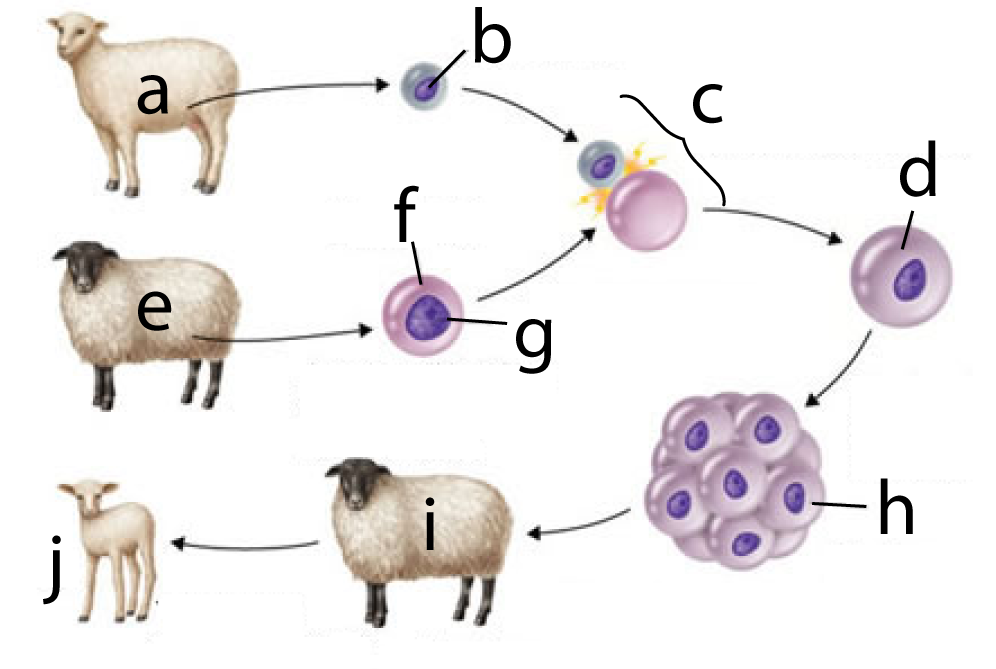
In 1996, Sir Ian Wilmut successfully cloned a mammal. To dramatize the outcome, he worked with two distinct breeds of sheep: a white-faced Finn Dorset (at “a”) and a Scottish Blackface (“e”). The process involved:
- Taking a cell from a female Finn Dorset’s mammary glands (“b”).
- Taking an unfertilized egg cell from a Scottish Blackface (“f”) and removing that cell’s nucleus (“g”).
- Fusing the mammary gland cell with the enucleated egg cell (at “c”), resulting in a diploid zygote (at “d”).
- The zygote grew into an embryo (h), which was then implanted into a surrogate mother (another Scottish blackface).
- When the lamb was born (“j”) it had a Finn Dorset phenotype, and subsequent genetic analysis showed that it was a clone of “a.”
Here’s a musical explanation of the process. For the lyrics, click here.
3. Cells Differentiate Because they Express Different Genes
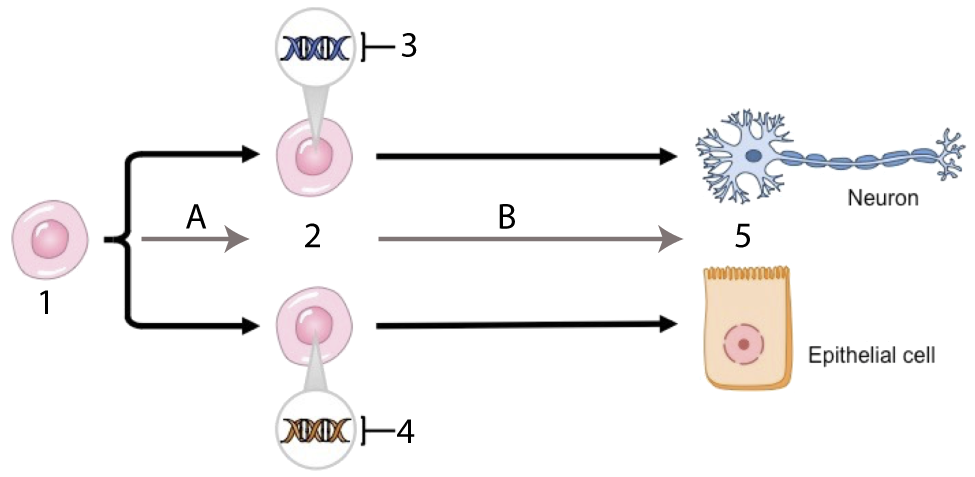
What’s the takeaway from reproductive cloning? It’s that all cells have the genetic information to create an entire organism. Why then, are cells of different tissue types so different?
Differences in cell structure and function are the result of differential gene expression. In the diagram at left, the neuron and epithelial cell are genomically equivalent. But in the undifferentiated cells (at 2) that gave rise to these mature cells, tissue-specific genes were activated. The genes that were activated at 3 were genes that produced neuron proteins; conversely, the genes activated at 4 were genes that produced epithelial cell proteins.
4. Checking Understanding: Genomic Equivalence, Cloning, and Differentiation
[qwiz random = “true” qrecord_id=”sciencemusicvideosMeister1961-Genomic Equivalence, Cloning, Differentiation (v2.0)”]
[h]Genomic Equivalence, Cloning, and Differentiation
[i]
[q] All cells are [hangman]equivalent.
[c]IGdlbm9taWNhbGx5[Qq]
[f]IEdvb2Qh[Qq]
[q] In this depiction, the genetic material of the organism John Gurdon wanted to be cloned is indicated by
[textentry single_char=”true”]
[c]ID M=[Qq]
[f]IEV4Y2VsbGVudC4gTnVtYmVyICYjODIyMDszJiM4MjIxOyBpcyB0aGUgbnVjbGV1cyBvZiBhIGNlbGwgZnJvbSB0aGUgYW5pbWFsIEd1cmRvbiB3YW50ZWQgdG8gY2xvbmUsIGFuZCBpdCBjb250YWlucyB0aGUgZ2VuZXRpYyBtYXRlcmlhbA==[Qq]
[c]IEVudGVyIHdvcmQ=[Qq]
[f]IFNvcnJ5LCB0aGF0JiM4MjE3O3Mgbm90IGNvcnJlY3Qu[Qq]
[c]ICo=[Qq]
[f]IE5vLiBMb29rIGZvciB0aGUgbnVjbGV1cyBmcm9tIGEgY2VsbCBmcm9tIHRoZSBkb25vciBvcmdhbmlzbS4=[Qq]
[q] In this depiction, an enucleated egg is indicated by
[textentry single_char=”true”]
[c]ID Y=[Qq]
[f]IEZhbnRhc3RpYy4gTnVtYmVyICYjODIyMDs2JiM4MjIxOyBpcyBhbiBlZ2cgY2VsbCB0aGF0IGhhcyBoYWQgaXRzIG51Y2xldXMgcmVtb3ZlZC4=[Qq]
[c]IEVudGVyIHdvcmQ=[Qq]
[f]IE5vLCB0aGF0JiM4MjE3O3Mgbm90IGNvcnJlY3Qu[Qq]
[c]ICo=[Qq]
[f]IE5vLiBMb29rIGZvciBhbiBlZ2cgY2VsbCB0aGF0IGhhcyBoYWQgaXRzIG51Y2xldXMgcmVtb3ZlZC4=[Qq]
[q] In terms of development, the key idea of the diagram below is that even a differentiated cell (like the one at “2”) has the [hangman] information to make an entirely new organism.
[c]IGdlbmV0aWM=[Qq]
[f]IEdvb2Qh[Qq]
[q] In the diagram below, a cloned embryo is shown at
[textentry single_char=”true”]
[c]IG g=[Qq]
[f]IEV4Y2VsbGVudC4gTGV0dGVyICYjODIyMDtoJiM4MjIxOyBzaG93cyBhIGNsb25lZCBlbWJyeW8u[Qq]
[c]IEVudGVyIHdvcmQ=[Qq]
[f]IE5vLCB0aGF0JiM4MjE3O3Mgbm90IGNvcnJlY3Qu[Qq]
[c]ICo=[Qq]
[f]IE5vLiBMb29rIGZvciB0aGUgZW1icnlvIHRoYXQgcmVzdWx0cyBmcm9tIHRoZSB6eWdvdGUgdGhhdCYjODIxNztzIGNyZWF0ZWQgd2hlbiB0aGUgZG9ub3IgY2VsbHMgaXMgZnVzZWQgd2l0aCB0aGUgZW51Y2xlYXRlZCBlZ2cu[Qq]
[q] In the diagram below, the surrogate mother is at
[textentry single_char=”true”]
[c]IG k=[Qq]
[f]IEV4Y2VsbGVudC4gJiM4MjIwO2kmIzgyMjE7IHNob3dzIHRoZSBzdXJyb2dhdGUgbW90aGVyLg==[Qq]
[c]IEVudGVyIHdvcmQ=[Qq]
[f]IE5vLg==[Qq]
[c]ICo=[Qq]
[f]IE5vLiBIZXJlJiM4MjE3O3MgYSBoaW50OsKgIEluIGNsb25pbmcgYSBtYW1tYWwsIHlvdSBuZWVkIGEgc3Vycm9nYXRlIG1vdGhlciB0byB0aGF0IHRoZSBlbWJyeW8gY2FuIGRldmVsb3AgdG8gbWF0dXJpdHku[Qq]
[q multiple_choice=”true”] In the neuron and the epithelial cell shown at number 5 below, which of the following would be the same?
[c]IFByb3RlaW4=[Qq]
[f]IE5vLiBUaGVzZSBjZWxscyBhcmUgZGlmZmVyZW50IGJlY2F1c2UgdGhleSYjODIxNztyZSB0cmFuc2xhdGluZyBkaWZmZXJlbnQgZ2VuZXMgaW50byBkaXN0aW5jdCBwcm90ZWlucy4=[Qq]
[c]IA==bVJOQQ==[Qq]
[f]Tm8uIFRoZXNlIGNlbGxzIGFyZSBkaWZmZXJlbnQgYmVjYXVzZSB0aGV5JiM4MjE3O3JlIGV4cHJlc3NpbmcgZGlmZmVyZW50IGdlbmVzLiBUaGVyZWZvcmUsIGRpZmZlcmVudCBnZW5lcyBhcmUgYmVpbmcgdHJhbnNjcmliZWQsIGxlYWRpbmcgdG8gdGhlIHByb2R1Y3Rpb24gb2YgZGlmZmVyZW50IG1STkFzLg==[Qq]
[c]IA ==RE5B[Qq]
[f]IFllcy4gQWxsIGNlbGxzIGFyZSBnZW5vbWljYWxseSBlcXVpdmFsZW50LCBzbyB5b3UgY291bGQgZXhwZWN0IHRoZXNlIHR3byBjZWxscywgZGVzcGl0ZSB0aGVpciBkaWZmZXJlbmNlcywgdG8gaGF2ZSB0aGUgc2FtZSBETkEu[Qq]
[q] In the diagram below, which letter represents differentiation?
[textentry single_char=”true”]
[c]IE I=[Qq]
[f]IE5pY2UhIExldHRlciAmIzgyMjA7QiYjODIyMTsgc2hvd3MgdGhlIHVuZGlmZmVyZW50aWF0ZWQgY2VsbHMgYXQgJiM4MjIwOzImIzgyMjE7IGNoYW5naW5nIGludG8gdGhlIGZ1bGx5IGRpZmZlcmVudGlhdGVkIGNlbGxzIGF0ICYjODIyMDs1LiYjODIyMTs=[Qq]
[c]IEVudGVyIHdvcmQ=[Qq]
[f]IE5vLg==[Qq]
[c]ICo=[Qq]
[f]IE5vLiBIZXJlJiM4MjE3O3MgYSBoaW50OsKgIFdoaWNoIGFycm93IGlzIHNob3dpbmcgdGhlIGNlbGxzIGNoYW5naW5nIGZyb20gYW4gdW5kaWZmZXJlbnRpYXRlZCBmb3JtICh3aGVyZSB0aGV5IGxvb2sgdGhlIHNhbWUpIHRvIGEgZGlmZmVyZW50aWF0ZWQgZm9ybSAod2hlcmUgdGhleSBsb29rIGRpZmZlcmVudD8p[Qq]
[/qwiz]
5. Differentiation can be caused by differences in the distribution of transcription factors in an egg
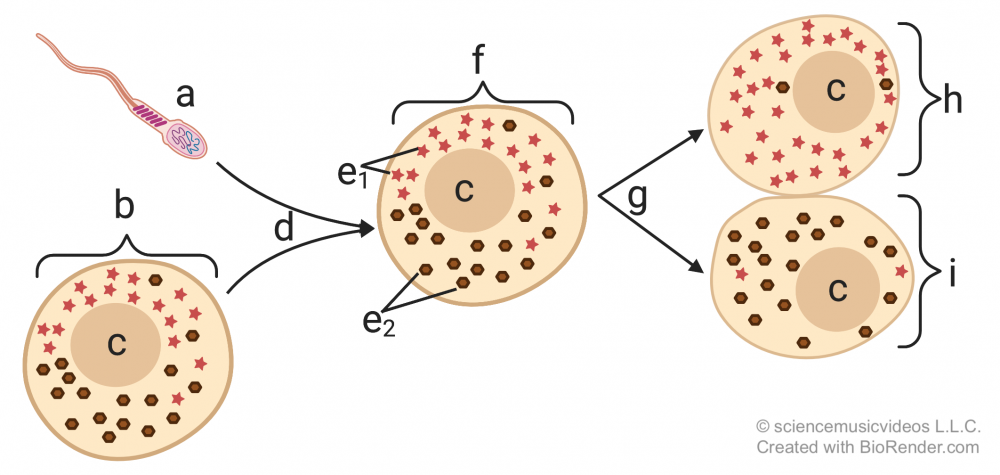
Differentiation starts early in the life of the organism— often in the very first cleavage divisions that follow fertilization. What causes the daughter cells to differentiate has to do with differences in the concentration of molecules in the cytoplasm that they receive from their parent.
In the diagram on the left, you can see this represented by the distribution of the molecules represented by the red star and the brown sphere (at “e”), with more of the red star molecules on the top side of the egg, and more of the brown sphere molecules on the bottom side. This uneven distribution is maintained after fertilization (“d”). As a result, when the zygote (“f”) divides (“g”), the result is two daughter cells with different concentrations of these molecules in their cytoplasm. If these molecules are proteins or RNAs that interact with the DNA in the daughter cells, they’re called cytoplasmic determinants, and the result will be the activation of different genes, creating the stage for the differentiation in the cell lines that stem from cells “h” and “i.”
 In the fruit fly Drosophila, an important model organism for developmental biology, one such cytoplasmic determinant is a type of mRNA named bicoid, which affects an embryo’s development of a head.
In the fruit fly Drosophila, an important model organism for developmental biology, one such cytoplasmic determinant is a type of mRNA named bicoid, which affects an embryo’s development of a head.
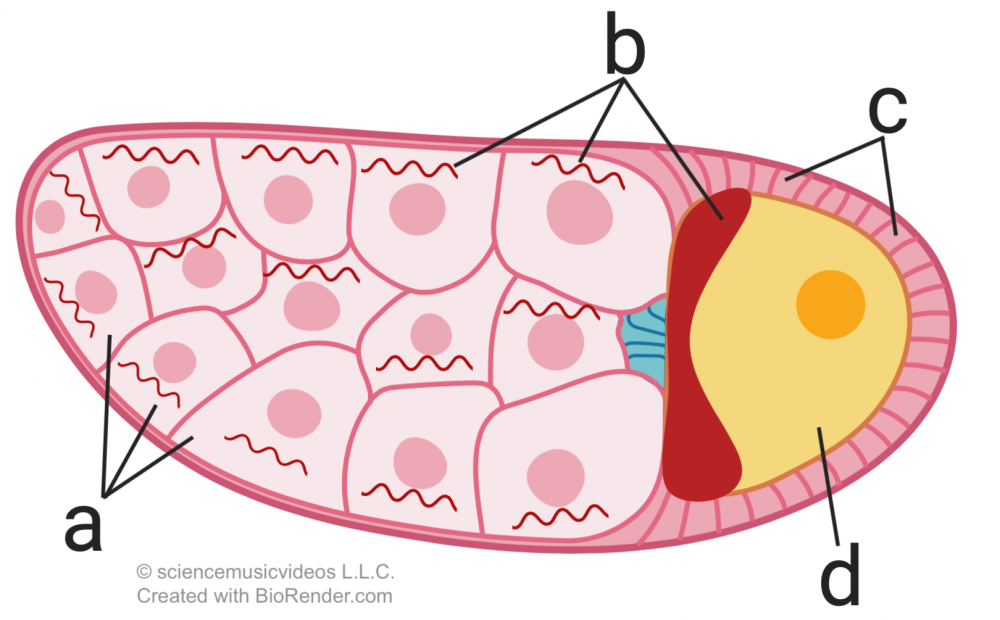 Here’s how bicoid works. Drosophila eggs (“d”), develop within a follicle (the entire structure shown on the right). At one end of the follicle are nurse cells (“a”). The nurse cells produce bicoid mRNA (“b”), which diffuses into the egg, and winds up being much more concentrated on one end of the egg cell than the other. Note that all of the cells except for “d” are diploid, maternal cells.
Here’s how bicoid works. Drosophila eggs (“d”), develop within a follicle (the entire structure shown on the right). At one end of the follicle are nurse cells (“a”). The nurse cells produce bicoid mRNA (“b”), which diffuses into the egg, and winds up being much more concentrated on one end of the egg cell than the other. Note that all of the cells except for “d” are diploid, maternal cells.
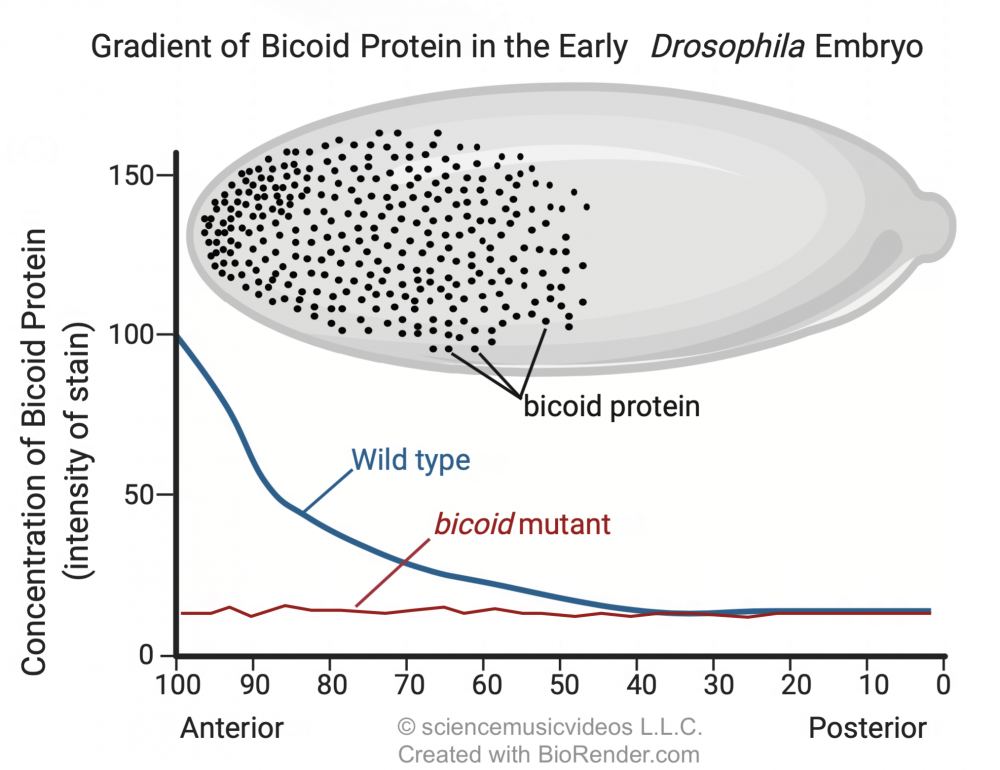
During development, the bicoid mRNA is translated into bicoid protein. The image below shows a gradient of bicoid protein in an early Drosophila embryo. The dark spots in the drawing show how the bicoid protein is concentrated in the head end of the developing embryo. The graph below the image quantifies this gradient in wild-type flies (see the blue line labeled “wild type”), and also in flies with mutant mothers that don’t express the bicoid gene (red line).
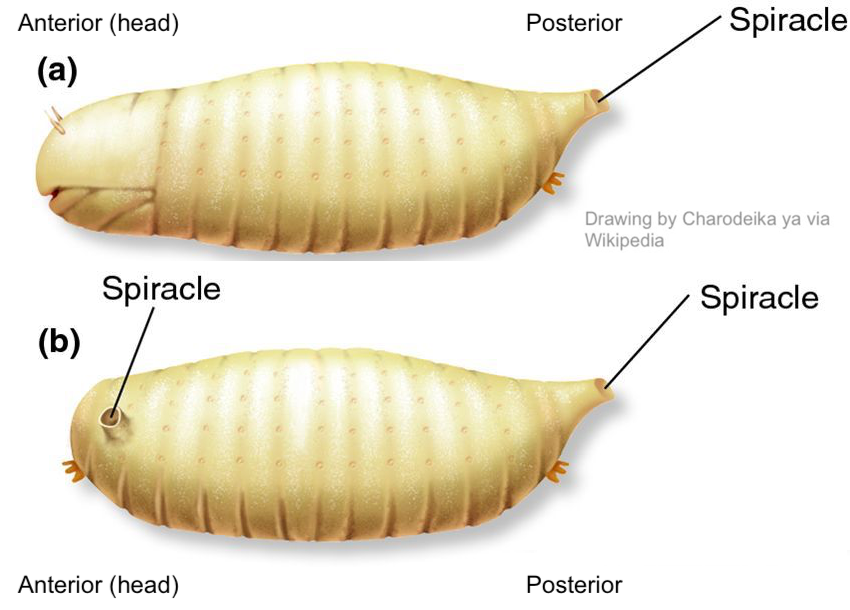
The presence of bicoid mRNA and bicoid protein causes a head to develop (shown at “a” below: note the mouth and antennae). Bicoid mutant mothers that don’t produce sufficient bicoid have headless offspring (shown at “b”). These quickly die.
What might be happening? The important move, as an AP Bio student, is to connect the presence or absence of bicoid protein to the regulatory mechanisms behind gene expression that you’ve learned about in previous tutorials. We know that the presence of bicoid is connected with cells on the head side of the embryo activating tissue-specific genes that lead to the differentiation of these tissues into a head. The specific mechanism might be at the level of DNA packaging. This could include
- Acetylation of tissue-specific genes in the head region, allowing them to be expressed, or
- Methylation of these same genes in the tail region, causing them not to be expressed.
Or the regulation could be at the level of transcription. In this case, bicoid protein could be acting as a transcription factor that turns on tissue-specific genes in the head region. With those transcription factors absent, a head can’t properly develop.
6. Differentiation can be caused by induction
 Cytoplasmic determinants like bicoid start the process of tissue differentiation that occurs during embryonic development. Another process connected with differentiation is induction.
Cytoplasmic determinants like bicoid start the process of tissue differentiation that occurs during embryonic development. Another process connected with differentiation is induction.
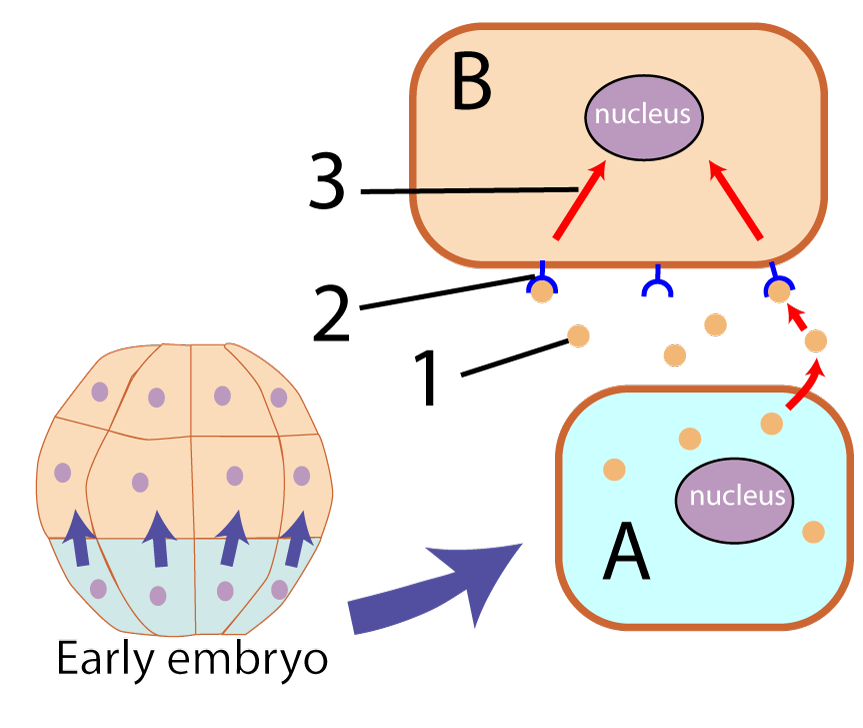
Induction involves one cell (or a group of cells) influencing the development of other cells. The inducing cells do this through the release of molecules that act as transcription factors. Such molecules, because they impact the form of the developing embryo, are often referred to as morphogens.
In the diagram on the left, the inducing cells are the ones shaded blue-green at the bottom of the developing embryo. These cells (one of which is shown at “A”), release morphogens (“1”) that bind with membrane receptors (“2”) on another cell (“B”). When the morphogenetic signal binds with the receptor, it sets off a signal transduction cascade that ultimately reaches the nucleus, activating (or deactivating) genes.
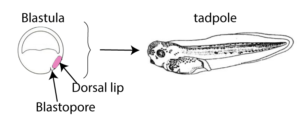
Graduate student Hilde Mangold, working with her advisor Otto Spemann, surgically removed the dorsal lip of the blastopore from an early embryo of a donor newt (shown in pink on the left embryo in the diagram below). They transplanted this tissue onto a second newt (the host, on the right) which now had two dorsal lips instead of one.
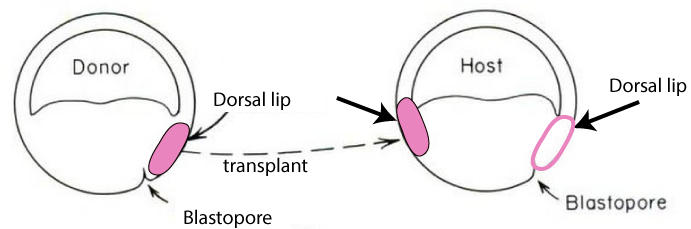
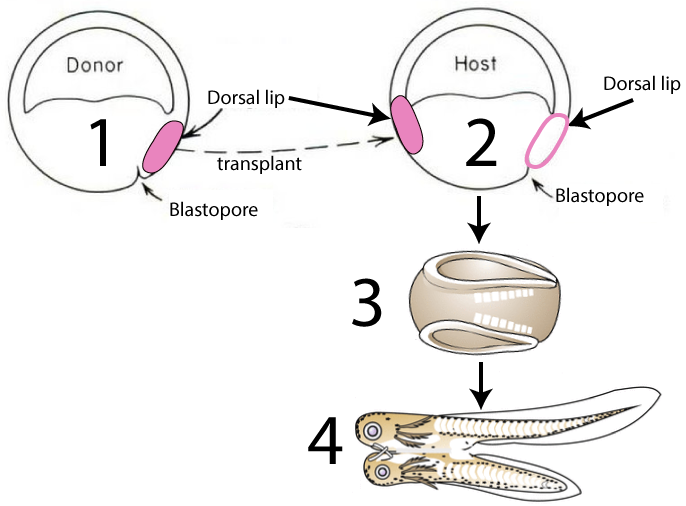
With two dorsal lips, the embryo developed into a conjoined newt (4).
7. Checking Understanding: Cytoplasmic Influences and Induction
[qwiz qrecord_id=”sciencemusicvideosMeister1961-Cytoplasmic Influences and Induction (v2.0)”]
[h]Quiz: Cytoplasmic Influences and Induction
[i]
[q] The diagram below shows that in a zygote, molecules in the cytoplasm are not distributed in a uniform way. Some of these molecules might be a type of early-acting morphogen called a cytoplasmic [hangman]. And if so, cells “h” and “i” will [hangman] into different types of cells.
[c]IGRldGVybWluYW50[Qq]
[f]IEdyZWF0IQ==[Qq]
[c]IGRpZmZlcmVudGlhdGU=[Qq]
[f]IEdvb2Qh[Qq]
[q] In the diagram below, the zygote is at
[textentry single_char=”true”]
[c]IG Y=[Qq]
[f]IEV4Y2VsbGVudCEgVGhlIHp5Z290ZSBpcyBhdCAmIzgyMjA7Zi4mIzgyMjE7[Qq]
[c]IEVudGVyIHdvcmQ=[Qq]
[f]IE5vLg==[Qq]
[c]ICo=[Qq]
[f]IE5vLiBIZXJlJiM4MjE3O3MgYSBoaW50OiBhIHNwZXJtIGNlbGwgaXMgYXQgJiM4MjIwO2EmIzgyMjE7LiBUaGUgenlnb3RlIGRldmVsb3BzIHdoZW4gdGhlIHNwZXJtIGFuZCBlZ2cgZnVzZSB0b2dldGhlciBkdXJpbmcgZmVydGlsaXphdGlvbi4=[Qq]
[q] In the diagram below, the first round of cell division is indicated by
[textentry single_char=”true”]
[c]IG c=[Qq]
[f]IE5pY2UhIFRoZSB6eWdvdGUmIzgyMTc7cyBmaXJzdCBmZXcgY2VsbCBkaXZpc2lvbnMgYXJlIHNob3duIGF0ICYjODIyMDtnLiYjODIyMTs=[Qq]
[c]IEVudGVyIHdvcmQ=[Qq]
[f]IE5vLg==[Qq]
[c]ICo=[Qq]
[f]IE5vLiBIZXJlJiM4MjE3O3MgYSBoaW50OiBMZXR0ZXIgJiM4MjIwO0MmIzgyMjE7IHJlcHJlc2VudHMgdGhlIHp5Z290ZS4gV2hhdCBwcm9jZXNzIGluZGljYXRlcyBpdHMgZGl2aXNpb24gaW50byB0d28gZGF1Z2h0ZXIgY2VsbHM/[Qq]
[q] If the diagram below is telling the story of cytoplasmic determinants, then the molecules indicated by “e” must be [hangman] factors
[c]IHRyYW5zY3JpcHRpb24=[Qq]
[f]IEV4Y2VsbGVudCE=[Qq]
[q] The diagram below shows a drosophila egg, surrounded by its follicle. What letter represents bicoid mRNA?
[textentry single_char=”true”]
[c]IG I=[Qq]
[f]IE5pY2UhIEJpY29pZCBtUk5BIGlzIHJlcHJlc2VudGVkIGJ5ICYjODIyMDtiLiYjODIyMTs=[Qq]
[c]IEVudGVyIHdvcmQ=[Qq]
[f]IE5vLg==[Qq]
[c]ICo=[Qq]
[f]IE5vLiBIZXJlJiM4MjE3O3MgYSBoaW50OiBtUk5BIGlzIGEgc2luZ2xlLXN0cmFuZGVkIG51Y2xlaWMgYWNpZC4gVGhpbmsgYWJvdXQgaG93IEROQSBpcyByZXByZXNlbnRlZCwgdGhlbiBpbWFnaW5lIGEgc2luZ2xlLXN0cmFuZGVkIHZlcnNpb24gb2YgdGhhdC4=[Qq]
[q multiple_choice=”true”] The headless fly shown at “b” below is caused by a genetic mutation in
[c]IHRoZSBjZWxscyBvZiB0aGUgZGV2ZWxvcGluZyBmbHkgZW1icnlv[Qq]
[f]IE5vLiBUaGUgZGV2ZWxvcGluZyBmbHkgZW1icnlvIG9ubHkgbmVlZHMgdG8gYmUgZXhwb3NlZCB0byBwcm9wZXIgbGV2ZWxzIG9mIGJpY29pZCBtUk5BIGFuZCBiaWNvaWQgcHJvdGVpbiB0byBkZXZlbG9wIGEgaGVhZC4gV2hlcmUgZG9lcyB0aGUgYmljb2lkIG1STkEgY29tZSBmcm9tPw==[Qq]
[c]IFRoZSBzcGVybSB0aGF0IGZlcnRpbGl6ZWQgdGhlIGVnZyBjZWxsLCBjcmVhdGluZyB0aGUgenlnb3Rl[Qq]
[f]IFRoZSBkZXZlbG9waW5nIGZseSBlbWJyeW8gb25seSBuZWVkcyB0byBiZSBleHBvc2VkIHRvIHByb3BlciBsZXZlbHMgb2YgYmljb2lkIG1STkEgYW5kIGJpY29pZCBwcm90ZWluIHRvIGRldmVsb3AgYSBoZWFkLiBXaGVyZSBkb2VzIHRoZSBiaWNvaWQgbVJOQSBjb21lIGZyb20/[Qq]
[c]IHRoZSBjZWxscyBvZi B0aGUgbW90aGVyLg==[Qq]
[f]IENvcnJlY3QhwqAgQSBtdXRhdGlvbiBpbiB0aGUgY2VsbHMgbWFraW5nIHVwIHRoZSBtb3RoZXImIzgyMTc7cyBlZ2cgZm9sbGljbGUgaXMgY2F1c2luZyBjZWxscyB0byBzZWNyZXRlIGluc3VmZmljaWVudCBiaWNvaWQgbVJOQSB0byBjb3JyZWN0bHkgZW5hYmxlIHRvIGRldmVsb3BpbmcgZW1icnlvIHRvIGNvcnJlY3RseSBidWlsZCBhIGhlYWQu[Qq]
[q] The image below is illustrating embryonic [hangman].
[c]IGluZHVjdGlvbg==[Qq]
[f]IEV4Y2VsbGVudCE=[Qq]
[q] In the system below, the inducing cells are shown at
[textentry single_char=”true”]
[c]IE E=[Qq]
[f]IEV4Y2VsbGVudC4gVGhlIGluZHVjaW5nIGNlbGxzIGFyZSBzaG93biBhdCAmIzgyMjA7QS4mIzgyMjE7[Qq]
[c]IEVudGVyIHdvcmQ=[Qq]
[f]IFNvcnJ5LCB0aGF0JiM4MjE3O3Mgbm90IGNvcnJlY3Qu[Qq]
[c]ICo=[Qq]
[f]IE5vLiBZb3UmIzgyMTc7cmUgbG9va2luZyBmb3IgdGhlIGNlbGwgdGhhdCYjODIxNztzIGVtaXR0aW5nIGEgc2lnbmFsLg==[Qq]
[q] If the system below is about induction, then a signal transduction cascade is being shown at …
[textentry single_char=”true”]
[c]ID M=[Qq]
[f]IEV4Y2VsbGVudC4gTnVtYmVyICYjODIyMDszJiM4MjIxOyByZXByZXNlbnRzIGEgc2lnbmFsIHRyYW5zZHVjdGlvbiBjYXNjYWRlLg==[Qq]
[c]IEVudGVyIHdvcmQ=[Qq]
[f]IE5vLCB0aGF0JiM4MjE3O3Mgbm90IGNvcnJlY3Qu[Qq]
[c]ICo=[Qq]
[f]IE5vLiBZb3UmIzgyMTc7cmUgbG9va2luZyBmb3Igc29tZXRoaW5nIGluIHRoZSByZWNlaXZpbmcgY2VsbCB0aGF0IHdvdWxkIHRyYW5zbWl0IHRoZSBtZXNzYWdlIGZyb20gdGhlIG1lbWJyYW5lIHRvIHRoZSBudWNsZXVzLg==[Qq]
[q multiple_choice=”true”] The main principle demonstrate by the experiment below is that
[c]IHRpc3N1ZXMgdGhhdCByZWxlYXNlIG1vcnBob2dlbnMgY2 FuIGluZHVjZSBjaGFuZ2VzIGluIG90aGVyIHRpc3N1ZXMu[Qq]
[f]RXhjZWxsZW50LiBUaGF0JiM4MjE3O3MgYSBncmVhdCBzdW1tYXJ5IG9mIHRoZSBTcGVtYW5uLU1hbmdvbGQgZXhwZXJpbWVudC4=[Qq]
[c]IGFsbCBjZWxscyBoYXZlIHRoZSBzYW1lIGdlbmVzOyB0aGV5IGp1c3QgZXhwcmVzcyBjZXJ0YWluIGdlbmVzLg==[Qq]
[f]Tm8uIFdoaWxlIHRoYXQmIzgyMTc7cyBhIHRydWUgc3RhdGVtZW50LCBpdCBkb2VzbiYjODIxNzt0IGNhcHR1cmUgd2hhdCYjODIxNztzIHNob3duIGFib3ZlLiBIZXJlJiM4MjE3O3MgYSBoaW50OiBOb3RpY2UgdGhlIGVmZmVjdCBvZiB0aGUgdHJhbnNwbGFudGVkIHRpc3N1ZSAoJiM4MjIwO2ImIzgyMjE7KS4=[Qq]
[q] In the diagram below, the transplanted tissue that was able to induce the formation of the second embryo was
[textentry single_char=”true”]
[c]IG I=[Qq]
[f]IE5pY2UuIExldHRlciAmIzgyMjA7YiYjODIyMTsgaXMgdGhlIHRyYW5zcGxhbnRlZCB0aXNzdWUgdGhhdCB3YXMgYWJsZSB0byBpbmR1Y2UgZGV2ZWxvcG1lbnQgb2YgYSBzZWNvbmQgZW1icnlvLg==[Qq]
[c]IEVudGVyIHdvcmQ=[Qq]
[f]IE5vLCB0aGF0JiM4MjE3O3Mgbm90IGNvcnJlY3Qu[Qq]
[c]ICo=[Qq]
[f]IE5vLiBZb3UmIzgyMTc7cmUgbG9va2luZyBmb3IgdGhlIHRyYW5zcGxhbnRlZCB0aXNzdWUgdGhhdCB3YXMgcmVzcG9uc2libGUgZm9yIGluZHVjaW5nIGEgbmV3IHJvdW5kIG9mIGVtYnJ5byBidWlsZGluZy4=[Qq]
[/qwiz]
8. Developmental genes are expressed in a hierarchical sequence, from general to specific
The images below show five vertebrate embryos at equivalent phases of development. The top row consists of photographs of the embryos, some of which were modified in order to highlight their similarities. For example, a large yolk sac was removed from the salamander embryo. The bottom row shows drawings from Ernst Haeckel, a famous 19th-century biologist.
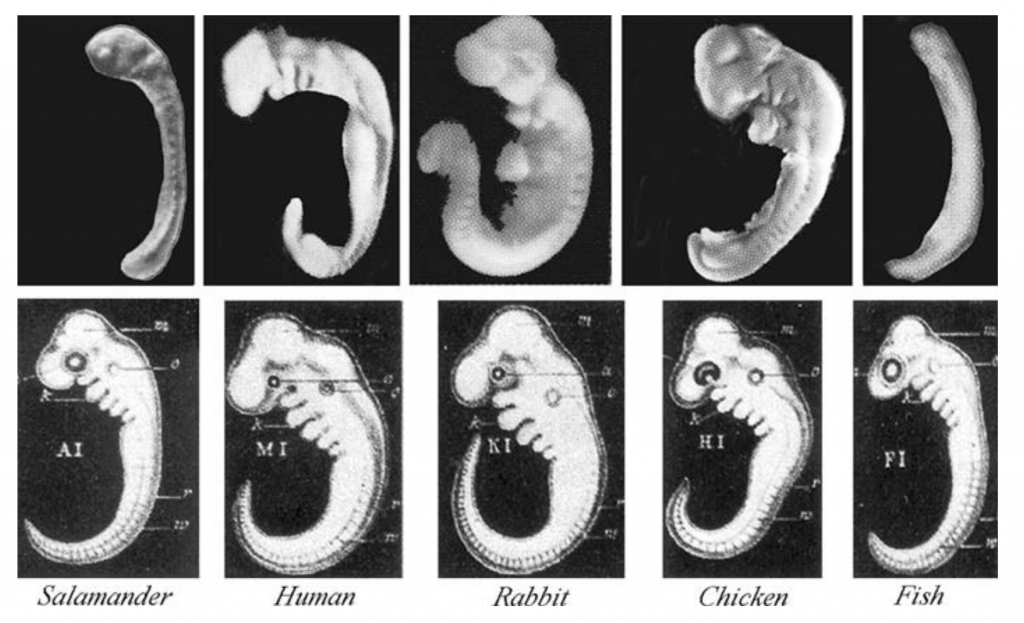
Haeckel created these drawings because he felt that they served as evidence for evolution, a topic that we’ll address in Unit 7 of our AP Biology course. For now, let’s use them to consider an important idea related to eukaryotic gene expression: the idea that developmental genes are expressed in a hierarchical sequence, from general to specific.
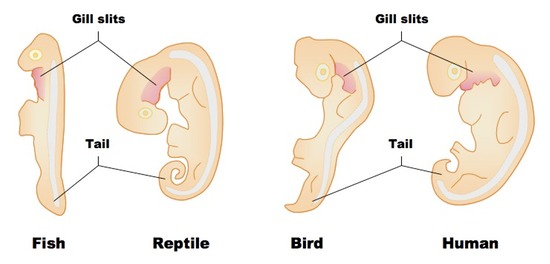
Start by focusing on the column showing a human embryo. The embryo shown here is only a few weeks old. At that moment in development, the embryo has a variety of traits that it will later lose: these include a tail and what are called pharyngeal slits (also known as gill slits: click on the images on the right and below for a better view).

Pharyngeal slits are openings that extend from the neck area below the head into the back of the mouth cavity. In a fish, these slits are maintained in the adult form. They let water flow over the gills, creating a current that enhances the absorption of oxygen.
In all tailless vertebrates, like humans and birds, the tail gets broken down through apoptosis: programmed cell death. The material making up the tail gets recycled and is used to build other parts of the body. The same thing happens to the gill slits in all of the four-limbed vertebrates. That’s because all of these animals (amphibians, reptiles, birds, and mammals) absorb oxygen from the air using their lungs, rather than using gills to absorb oxygen from the water.
Why do the embryos of humans, rabbits, and chickens develop with tails and gill slits, only to break them down and absorb them later? It’s because having a tail (for example) is a part of a general developmental plan inherited from a vertebrate ancestor. In early embryonic forms, the genes for this plan are still expressed. The only reason our embryonic form possesses a tail is because a tail is part of a developmental program that we inherited from our ancestors.
Why haven’t the genes for these structures been deleted? We’ll focus on the tail. One possibility is that the developmental plan to build a 4-limbed vertebrate might require that at some moment in development, a tail be present. Without having an embryonic tail, the complex signaling that goes on between cells which results in the various parts of the body growing in the right place could be disrupted. So all vertebrates start life by building a tail, whether they keep it or not. The same is true of the gill slits.
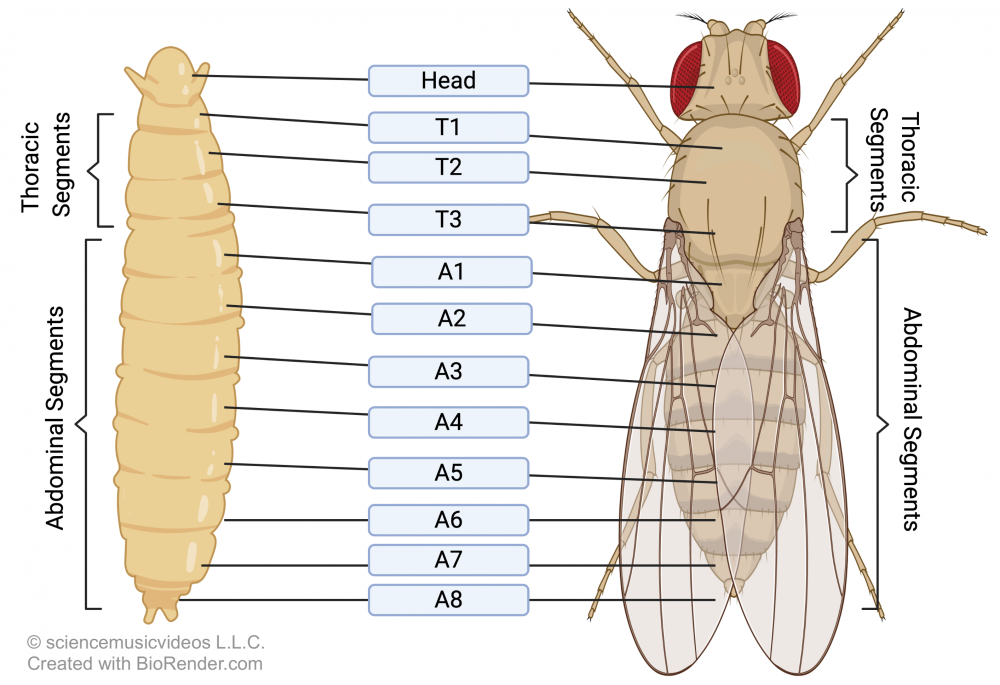
Expression of genes in a hierarchical pattern, with genes for general features laid down before genes for specific features, can also be seen in the emergence of segmentation in arthropods like Drosophila.
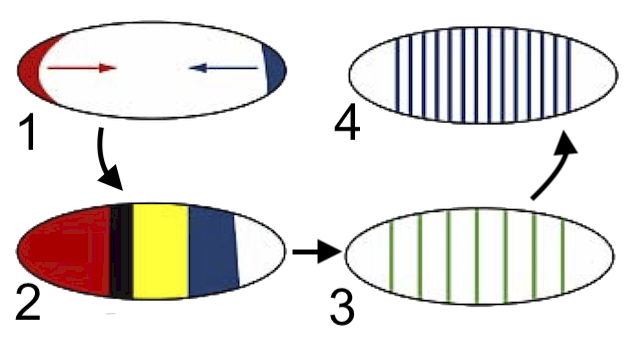
As a Drosophila embryo develops, among the first genes expressed are those that lay down the head-to-tail axis (1). The next set of genes to be activated divide the embryo into a few major regions (2). The next divide these regions into specific segments (3 and 4), each of which has an anterior end and a posterior end.
Only once these segments have been established are additional genes expressed, leading to the emergence of specific appendages on each segment.
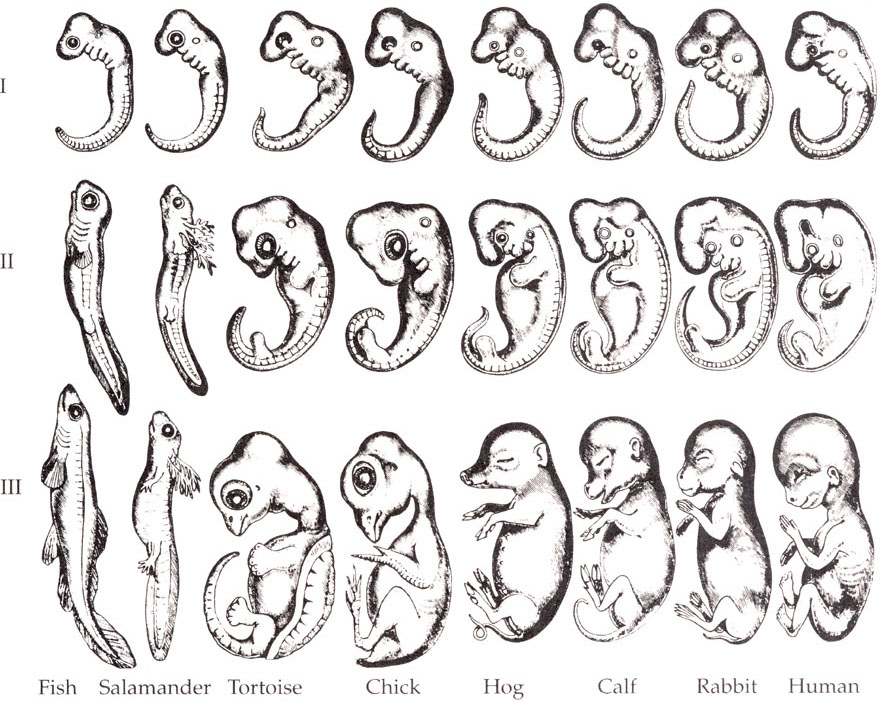
We’ll end by looking once again at Haekel’s drawings about embryological development in vertebrates. What’s the takeaway? The key idea is that during development, gene activation and induction unfold in a hierarchical sequence. The most general features (in this case, the basic vertebrate body plan shown in row 1) are laid down first. In rows II and three, genes that are specific to each species are expressed, with the embryos becoming more and more differentiated as development proceeds.
How does this happen? Through all of the mechanisms we’ve seen in this and previous tutorials for controlling gene expressions. To review, these include:
- epigenetic modifications of DNA through chromatin packaging,
- control of transcription through transcription factors,
- post-transcriptional modifications of RNA,
- regulatory RNAs such as micro RNA.
- cytoplasmic determinants in the egg
- induction.
9. Eukaryotic Gene Expression and Development Flashcards
[qdeck bold_text=”false” style=”min-height: 400px !important; width: 550px !important;” qrecord_id=”sciencemusicvideosMeister1961-Gene expression and Development FC (v2.0)”]
[h] Gene expression and Development Flashcards
[q] Using the diagram below, explain what cloning is, and explain how a mammal can be cloned.
[a] Cloning is a type of asexual reproduction in which a genetic copy is made of an organism. It involves extracting a nucleus from a somatic cell (b) and inserting it into an enucleated egg (c). The resulting zygote develops into an embryo which is transplanted into a surrogate mother (i). The clone is shown at “j.”
[q] In terms of eukaryotic gene expression, what is the important principle shown by the diagram below?
[a] The idea shown below is genomic equivalence. This is the idea that (with a few exceptions like gametes) all cells in a multicellular eukaryotic organism have the same DNA. The reason why different tissues are different is that they express different genes.
[q] Using the diagram below as a stimulus, explain the role of cytoplasmic determinants in cellular differentiation in multicellular eukaryotes.
[a] The diagram below is showing how the initial difference in the distribution of molecules in the cytoplasm of the egg can set the stage for subsequent tissue differentiation. If the molecules (represented by e1 and e2 below) are cytoplasmic determinants —proteins that act as transcription factors in the daughter cells — the result will be the activation of different genes, creating the stage for the differentiation in the cell lines that stem from cells “h” and “i.”
[q] Bicoid is an example of a cytoplasmic determinant that affects head development. Using the diagram below, explain how bicoid works.
[a] Drosophila eggs form within a follicle. The follicle cells secrete Bicoid mRNA, which accumulates in the head end of a Drosophila zygote. During development, the bicoid mRNA is translated into bicoid protein, which, like bicoid mRNA, is more concentrated in the head side of the embryo than it is on the tail side. As a result, the embryo develops a proper head (a). Bicoid mutant mothers don’t produce bicoid mRNA. As a result, they produce headless embryos (b), which quickly die.
[q] What is induction?
[a] Induction involves one group of cells producing molecules that induce developmental changes in other cells. In the diagram below, cell “A” is the inducing cell. It sends a signal (called a morphogen, at “1”) to another cell (B). Reception of the morphogen causes a signaling cascade (3) which activates genes in the receiving cell.
[q] Human embryos (along with those of birds and reptiles) possess gill slits and tails. How can this be explained within the context of eukaryotic gene expression?
[a] During development, processes such as gene activation and induction unfold in a hierarchical sequence. Genes for expressing general features are expressed first. Then genes for more specific features are expressed. In the example above, genes for the general vertebrate body plan are expressed first, producing an embryo that has gill slits and a tail. Later, genes for land-living vertebrates (with lungs) are expressed, and the gill slits tissue is absorbed and repurposed for other structures and functions. In humans and other primates without a tail, the tail tissue is absorbed.
[q]Connect the diagram below to the idea that embryonic development involves sequential gene expression..
[a] During embryonic development, genes are expressed in a way in which a general form is expressed before a more specific form. For example, in the development of the fruit fly Drosophila melanogaster, among the first genes expressed are those that lay down the head-to-tail axis (1). The next set of genes to be activated divide the embryo into a few major regions (2). The next divide these regions into specific segments (3 and 4), each of which has an anterior end and a posterior end. Finally, genes are expressed, determining which appendages are expressed on each segment.
[x] [restart]
[/qdeck]
10. What’s Next?
Proceed to Topics 6.5 – 6.6, Part 6: Eukaryotic gene regulation and evolutionary change in the threespine stickleback (the next tutorial in AP Bio Unit 6).